, Jurkat (2), Hela (3), HEK293 (4) and A549 (5) cell lysate.")
 and kidney cancer tissues (right) using SIRT1 mouse mAb with DAB staining.")
. Red: Actin filaments have been labeled with Alexa Fluor-555 phalloidin.")
 The effects of glucose concentration (25, 30, 35, 40, 45, 50, 55, 60, 70, 80 and 90 mM) on viability of HepG2 cells by MTT. (B) The effects of induced time (0, 12, 24, 48, 72 and 96 h) exposed to 60 mM glucose on the viability of HepG2 cells by MTT. (C) The TG and T-CHO levels in HepG2 cells exposed to 25 mM, 45 mM and 60 mM glucose for 48 h. (D) The TG and T-CHO levels in HepG2 cells exposed to 60 mM glucose for 0 h, 24 h and 48 h. (E) Oil Red O staining in different groups of HepG2 cells. (F, G) The relative protein levels and distribution of Sirt1 in HepG2 cells exposed to 25 mM, 45 mM and 60 mM glucose for 48 h. (H, I) The relative protein levels and distribution of Sirt1 in HepG2 cells exposed to 60 mM glucose for 0 h, 24 h and 48 h. Each bar represents the mean ± SEM for groups of three. *P < 0.05, **P < 0.01, compared to 0 h or 25 mM as indicated.")
 The effects of glucose concentration (25, 30, 35, 40, 45, 50, 55, 60, 70, 80 and 90 mM) on viability of HepG2 cells by MTT. (B) The effects of induced time (0, 12, 24, 48, 72 and 96 h) exposed to 60 mM glucose on the viability of HepG2 cells by MTT. (C) The TG and T-CHO levels in HepG2 cells exposed to 25 mM, 45 mM and 60 mM glucose for 48 h. (D) The TG and T-CHO levels in HepG2 cells exposed to 60 mM glucose for 0 h, 24 h and 48 h. (E) Oil Red O staining in different groups of HepG2 cells. (F, G) The relative protein levels and distribution of Sirt1 in HepG2 cells exposed to 25 mM, 45 mM and 60 mM glucose for 48 h. (H, I) The relative protein levels and distribution of Sirt1 in HepG2 cells exposed to 60 mM glucose for 0 h, 24 h and 48 h. Each bar represents the mean ± SEM for groups of three. *P < 0.05, **P < 0.01, compared to 0 h or 25 mM as indicated.")
 SIRT1/FOXO1 pathway protein expression in MCAO/R rats; (B-D) Expression of SIRT1, FOXO1 and Ac-FOXO1 proteins in MCAO/R rats; (E) Protein expression of SIRT1/FOXO1 pathway in OGD/R PC12 cells; (F-H) Expression of SIRT1, FOXO1 and Ac-FOXO1 proteins in OGD/R PC12 cells. (n = 3, #P < 0.05 vs Sham; ##P < 0.01 vs Sham or Control; *P < 0.05 vs Model or OGD/R; **P < 0.01 vs Model or OGD/R).")
 Protein interaction analysis. (B) Expression levels of SIRT1 and MFN2 proteins in the heart tissues of mice. (C) Representative immunohistochemistry images showing the expression of SIRT1 and MFN2 in myocardial tissue. The arrows indicate areas of stained cells. Scale bar, 100 µm; magnification, ×40. (D) SIRT1 and MFN 2 were detected by immunofluorescence double staining to assess colocalization. Scale bars, 100 and 50 µm; magnification, ×40. Data are presented as the mean ± standard error of the mean (n=3); statistical analysis was performed using one-way ANOVA followed by Tukey's post hoc test. *P")
产品描述
*The optimal dilutions should be determined by the end user. For optimal experimental results, antibody reuse is not recommended.
*Tips:
WB: 适用于变性蛋白样本的免疫印迹检测. IHC: 适用于组织样本的石蜡(IHC-p)或冰冻(IHC-f)切片样本的免疫组化/荧光检测. IF/ICC: 适用于细胞样本的荧光检测. ELISA(peptide): 适用于抗原肽的ELISA检测.
引用格式: Affinity Biosciences Cat# BF0189, RRID:AB_2833834.
展开/折叠
75SirT1; hSIR2; hSIRT1; HST2, S. cerevisiae, homolog of; NAD dependent deacetylase sirtuin 1; NAD dependent protein deacetylase sirtuin 1; OTTHUMP00000198111; OTTHUMP00000198112; Regulatory protein SIR2 homolog 1; SIR1_HUMAN; SIR2; SIR2 like 1; SIR2 like protein 1; SIR2, S.cerevisiae, homolog-like 1; SIR2-like protein 1; SIR2ALPHA; SIR2L1; Sirt1; SirtT1 75 kDa fragment; Sirtuin (silent mating type information regulation 2 homolog) 1 (S. cerevisiae); Sirtuin 1; Sirtuin type 1;
抗原和靶标
Purified recombinant fragment of human SIRT1 expressed in E. Coli.
研究领域
· Cellular Processes > Cell growth and death > Cellular senescence. (View pathway)
· Environmental Information Processing > Signal transduction > FoxO signaling pathway. (View pathway)
· Environmental Information Processing > Signal transduction > AMPK signaling pathway. (View pathway)
· Human Diseases > Substance dependence > Amphetamine addiction.
· Human Diseases > Cancers: Overview > MicroRNAs in cancer.
· Organismal Systems > Aging > Longevity regulating pathway. (View pathway)
· Organismal Systems > Aging > Longevity regulating pathway - multiple species. (View pathway)
· Organismal Systems > Endocrine system > Glucagon signaling pathway.
文献引用
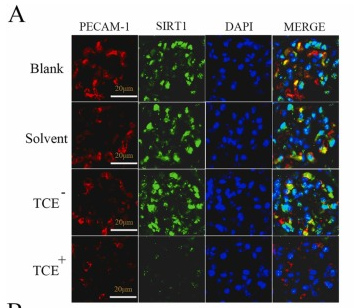
Application: IF/ICC Species: Mouse Sample: endothelial cells

Application: WB Species: Rat Sample:

Application: WB Species: Mouse Sample: liver tissues

Application: IF/ICC Species: Mouse Sample: liver tissues

Application: WB Species: Mouse Sample:
限制条款
产品的规格、报价、验证数据请以官网为准,官网链接:www.affbiotech.com | www.affbiotech.cn(简体中文)| www.affbiotech.jp(日本語)产品的数据信息为Affinity所有,未经授权不得收集Affinity官网数据或资料用于商业用途,对抄袭产品数据的行为我们将保留诉诸法律的权利。
产品相关数据会因产品批次、产品检测情况随时调整,如您已订购该产品,请以订购时随货说明书为准,否则请以官网内容为准,官网内容有改动时恕不另行通知。
Affinity保证所销售产品均经过严格质量检测。如您购买的商品在规定时间内出现问题需要售后时,请您在Affinity官方渠道提交售后申请。产品仅供科学研究使用。不用于诊断和治疗。
产品未经授权不得转售。
Affinity Biosciences将不会对在使用我们的产品时可能发生的专利侵权或其他侵权行为负责。Affinity Biosciences, Affinity Biosciences标志和所有其他商标所有权归Affinity Biosciences LTD.